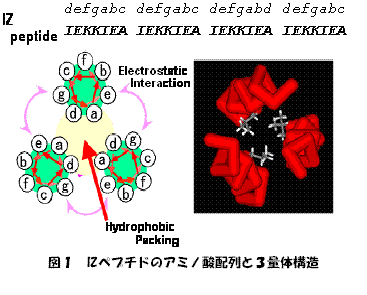
 僐僀儖僪僐僀儖峔憿偼丄偼偠傔僩儘億儈僆僔儞偺傛偆側慇堐忬抈敀幙偺峔惉惉暘偲偟偰揤慠抈敀幙偺攝楍偺拞偐傜尒偄偩偝傟偰偄偨偑丄傗偑偰丄1988擭偵S. L. Mcknight丄R. Tjian傜偵傛傝揤慠偺揮幨場巕抈敀幙偲偟偰傛偔抦傜傟傞GCN4, Fos, Jun, Myc摍偺拞偵尒傜傟傞Leucine Zipper峔憿偲偟偰抦傜傟傞傛偆偵側偭偨丅挿偝偵傕傛傞偑丄婎杮揑偵偼悈梟惈偺夛崌惈抈敀幙偱偁傝丄慳悈応偵娷傑傟傞Leu丄Ile摍偺慳悈惈巆婎娫偺慳悈惈憡屳嶌梡偲僿儕僢僋僗娫偺奅柺偵偍偗傞惷揹憡屳嶌梡側偳傪嬱摦椡偵丄俀偐傜俆杮偺塃姫偒偺兛亅僿儕僢僋僗偑嵍姫偒偵懇側偭偨丄僗乕僷乕僐僀儖峔憿傪庢傞丅俀検懱宆僐僀儖僪僐僀儖偼Leucine Zipper峔憿偲偟偰峀偔抦傜傟偰偄傞偑丄俁検懱宆偵偮偄偰偼揤慠偵偼儅僋儘僼傽乕僕僗僇儀儞僕儍乕儗僙僾僞乕丄僶僋僥儕僆僼傽乕僕偺gp41丄僸乕僩僔儑僢僋揮幨場巕側偳偵偍偄偰抦傜傟偰偄傞丅偙傟傜偺揤慠攝楍偼丄僐僀儖僪僐僀儖峔憿偺宍惉偵廳梫側慡偰偺憡屳嶌梡偑岠壥揑偵摥偔傛偆偵傾儈僲巁巆婎偑攝抲偝傟偰偄傞栿偱偼側偄堊丄儔僔儑僫儖側嵟揔壔偐傜Isoleucine Zipper乮IZ乯儁僾僠僪傪僆儕僕僫儖偵僨僓僀儞偟偨(恾侾)丅偙偺儁僾僠僪偼旕忢偵埨掕偱偁傝丄梈夝壏搙偼俋俆亷傪挻偊傞丅
僐僀儖僪僐僀儖峔憿偼丄偼偠傔僩儘億儈僆僔儞偺傛偆側慇堐忬抈敀幙偺峔惉惉暘偲偟偰揤慠抈敀幙偺攝楍偺拞偐傜尒偄偩偝傟偰偄偨偑丄傗偑偰丄1988擭偵S. L. Mcknight丄R. Tjian傜偵傛傝揤慠偺揮幨場巕抈敀幙偲偟偰傛偔抦傜傟傞GCN4, Fos, Jun, Myc摍偺拞偵尒傜傟傞Leucine Zipper峔憿偲偟偰抦傜傟傞傛偆偵側偭偨丅挿偝偵傕傛傞偑丄婎杮揑偵偼悈梟惈偺夛崌惈抈敀幙偱偁傝丄慳悈応偵娷傑傟傞Leu丄Ile摍偺慳悈惈巆婎娫偺慳悈惈憡屳嶌梡偲僿儕僢僋僗娫偺奅柺偵偍偗傞惷揹憡屳嶌梡側偳傪嬱摦椡偵丄俀偐傜俆杮偺塃姫偒偺兛亅僿儕僢僋僗偑嵍姫偒偵懇側偭偨丄僗乕僷乕僐僀儖峔憿傪庢傞丅俀検懱宆僐僀儖僪僐僀儖偼Leucine Zipper峔憿偲偟偰峀偔抦傜傟偰偄傞偑丄俁検懱宆偵偮偄偰偼揤慠偵偼儅僋儘僼傽乕僕僗僇儀儞僕儍乕儗僙僾僞乕丄僶僋僥儕僆僼傽乕僕偺gp41丄僸乕僩僔儑僢僋揮幨場巕側偳偵偍偄偰抦傜傟偰偄傞丅偙傟傜偺揤慠攝楍偼丄僐僀儖僪僐僀儖峔憿偺宍惉偵廳梫側慡偰偺憡屳嶌梡偑岠壥揑偵摥偔傛偆偵傾儈僲巁巆婎偑攝抲偝傟偰偄傞栿偱偼側偄堊丄儔僔儑僫儖側嵟揔壔偐傜Isoleucine Zipper乮IZ乯儁僾僠僪傪僆儕僕僫儖偵僨僓僀儞偟偨(恾侾)丅偙偺儁僾僠僪偼旕忢偵埨掕偱偁傝丄梈夝壏搙偼俋俆亷傪挻偊傞丅
丂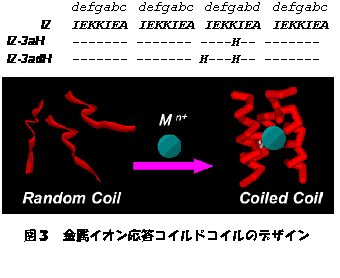 揤慠偺僐僀儖僪僐僀儖抈敀幙偵偍偄偰偼丄枾側慳悈憡屳嶌梡偑慳悈応偱宍惉偝傟偰偍傝丄堦斒偵梟攠丄僀僆儞側偳偼拞偵庢傝崬傑傟傞偙偲偼側偄丅偙傟偼丄慳悈応偵墬偗傞枾側慳悈僷僢僉儞僌偑僐僀儖僪僐僀儖宍惉偵偍偄偰旕忢偵廳梫偱偁傞帠傪帵偟偰偄傞丅偟偐偟側偑傜偙傟傪棤曉偣偽丄偙偺慳悈応傪巪偔僨僓僀儞偱偒傟偽丄嬐偐側憡屳嶌梡偵傛傝丄抈敀幙峔憿偺寑揑側曄壔乮僗僀僢僠乯偑壜擻偲側傞偙偲偑婜懸偱偒傞丅偦偙偱変乆偼偙偺慳悈応偵拲栚偟丄偄偔偮偐偺Ile傪His偵抲偒姺偊傞帠偱丄嬥懏攝埵擻傪帩偨偣傞帠傪帋傒偨丅嬥懏僀僆儞晄嵼壓偵偍偄偰偼丄慳悈僷僢僉儞僌偑曵傟傞帠偵傛傝僐僀儖僪僐僀儖峔憿偼宍惉偱偒偢乮儔儞僟儉峔憿乯丄嬥懏僀僆儞攝埵屻偼丄嶖宍惉偵敽偆僄僱儖僊乕偺棙摼偵傛傝僐僀儖僪僐僀儖峔憿傪宍惉偱偒傞帠傪婜懸偟偨丅僨僓僀儞偟偨攝楍偼恾俁偵帵偟偨丅慳悈応偺Ile傪侾偮His偵曄堎傪偐偗偨IZ-3aH偼丄嬥懏僀僆儞埶懚揑偵僐僀儖僪僐僀儖峔憿偑宍惉偝傟傞帠偑帵偝傟偨丅偙偺嵺偺慖戰惈偼丄Cu2+>Zn2+>>Ni2+偱偁偭偨丅堦曽偱俀偮His偵曄堎傪偐偗偨IZ-3adH偵偮偄偰傕嬥懏僀僆儞埶懚揑側僐僀儖僪僐僀儖峔憿偺宍惉偑娤應偝傟偨偑丄慖戰惈偼Ni2+乣Cu2+>>Zn2+偱偁偭偨丅傑偨丄偙偺俀偮偼巪偔堦偮偺抈敀幙偵撪偵慻傒崌傢偣傞帠偵傛傝丄堎庬嬥懏傪攝埵偱偒傞僐僀儖僪僐僀儖抈敀幙偺僨僓僀儞傕壜擻偱偁傞帠偑暘偐偭偨丅
揤慠偺僐僀儖僪僐僀儖抈敀幙偵偍偄偰偼丄枾側慳悈憡屳嶌梡偑慳悈応偱宍惉偝傟偰偍傝丄堦斒偵梟攠丄僀僆儞側偳偼拞偵庢傝崬傑傟傞偙偲偼側偄丅偙傟偼丄慳悈応偵墬偗傞枾側慳悈僷僢僉儞僌偑僐僀儖僪僐僀儖宍惉偵偍偄偰旕忢偵廳梫偱偁傞帠傪帵偟偰偄傞丅偟偐偟側偑傜偙傟傪棤曉偣偽丄偙偺慳悈応傪巪偔僨僓僀儞偱偒傟偽丄嬐偐側憡屳嶌梡偵傛傝丄抈敀幙峔憿偺寑揑側曄壔乮僗僀僢僠乯偑壜擻偲側傞偙偲偑婜懸偱偒傞丅偦偙偱変乆偼偙偺慳悈応偵拲栚偟丄偄偔偮偐偺Ile傪His偵抲偒姺偊傞帠偱丄嬥懏攝埵擻傪帩偨偣傞帠傪帋傒偨丅嬥懏僀僆儞晄嵼壓偵偍偄偰偼丄慳悈僷僢僉儞僌偑曵傟傞帠偵傛傝僐僀儖僪僐僀儖峔憿偼宍惉偱偒偢乮儔儞僟儉峔憿乯丄嬥懏僀僆儞攝埵屻偼丄嶖宍惉偵敽偆僄僱儖僊乕偺棙摼偵傛傝僐僀儖僪僐僀儖峔憿傪宍惉偱偒傞帠傪婜懸偟偨丅僨僓僀儞偟偨攝楍偼恾俁偵帵偟偨丅慳悈応偺Ile傪侾偮His偵曄堎傪偐偗偨IZ-3aH偼丄嬥懏僀僆儞埶懚揑偵僐僀儖僪僐僀儖峔憿偑宍惉偝傟傞帠偑帵偝傟偨丅偙偺嵺偺慖戰惈偼丄Cu2+>Zn2+>>Ni2+偱偁偭偨丅堦曽偱俀偮His偵曄堎傪偐偗偨IZ-3adH偵偮偄偰傕嬥懏僀僆儞埶懚揑側僐僀儖僪僐僀儖峔憿偺宍惉偑娤應偝傟偨偑丄慖戰惈偼Ni2+乣Cu2+>>Zn2+偱偁偭偨丅傑偨丄偙偺俀偮偼巪偔堦偮偺抈敀幙偵撪偵慻傒崌傢偣傞帠偵傛傝丄堎庬嬥懏傪攝埵偱偒傞僐僀儖僪僐僀儖抈敀幙偺僨僓僀儞傕壜擻偱偁傞帠偑暘偐偭偨丅
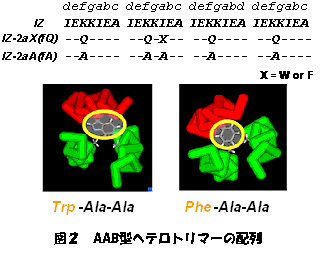
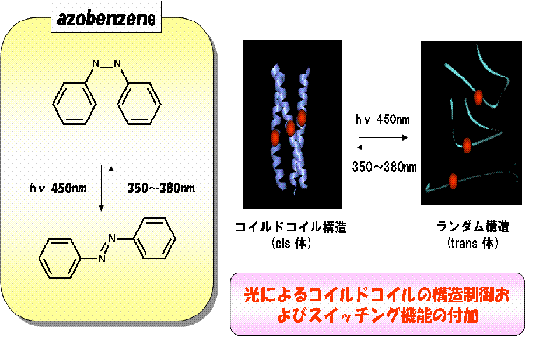 丂嵶朎撪摍偺娐嫬偱敪尰偝偣偰棙梡偡傞偙偲傪峫偊傞側傜丄旕揤慠桳婡壔崌暔傪慻傒崌傢偣偨恖岺抈敀幙偺愝寁偼堄枴傪側偝側偄偑丄嬤擭嵶朎撪傊偺條乆側抈敀幙摫擖庤朄偺専摙偑惉偝傟偰偄傞偙偲偐傜丄応柺偵傛偭偰偼桳岠側庤朄偲側傞偙偲偑婜懸偱偒傞丅偙偙偱偼丄俁検懱宆僐僀儖僪僐僀儖傊偺傾僝儀儞僛儞桿摫懱偺摫擖偵傛傞峔憿曄堎偵偮偄偰専摙傪峴偭偨丅梡偄偨傾僝儀儞僛儞桿摫懱偼丄
丂嵶朎撪摍偺娐嫬偱敪尰偝偣偰棙梡偡傞偙偲傪峫偊傞側傜丄旕揤慠桳婡壔崌暔傪慻傒崌傢偣偨恖岺抈敀幙偺愝寁偼堄枴傪側偝側偄偑丄嬤擭嵶朎撪傊偺條乆側抈敀幙摫擖庤朄偺専摙偑惉偝傟偰偄傞偙偲偐傜丄応柺偵傛偭偰偼桳岠側庤朄偲側傞偙偲偑婜懸偱偒傞丅偙偙偱偼丄俁検懱宆僐僀儖僪僐僀儖傊偺傾僝儀儞僛儞桿摫懱偺摫擖偵傛傞峔憿曄堎偵偮偄偰専摙傪峴偭偨丅梡偄偨傾僝儀儞僛儞桿摫懱偼丄